Cells Operate in an Order Fashion Under the Influence of
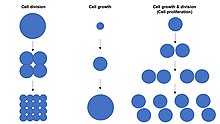
Jail cell division, growth & proliferation
Prison cell growth refers to an increase in the total mass of a cell, including both cytoplasmic, nuclear and organelle book.[1] Prison cell growth occurs when the overall charge per unit of cellular biosynthesis (production of biomolecules or anabolism) is greater than the overall charge per unit of cellular degradation (the destruction of biomolecules via the proteasome, lysosome or autophagy, or catabolism).[2] [3] [4]
Cell growth is not to be confused with cell division or the jail cell cycle, which are distinct processes that can occur alongside cell growth during the process of jail cell proliferation, where a prison cell, known as the "mother cell", grows and divides to produce ii "daughter cells".[1] Importantly, jail cell growth and cell partition can also occur independently of one another. During early embryonic development (cleavage of the zygote to form a morula and blastoderm), jail cell divisions occur repeatedly without cell growth. Conversely, some cells can abound without jail cell partition or without whatsoever progression of the cell cycle, such every bit growth of neurons during axonal pathfinding in nervous system evolution.
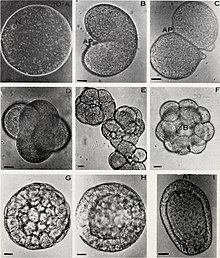
Cell division without cell growth during embryonic cleavage
In multicellular organisms, tissue growth rarely occurs solely through cell growth without cell sectionalization, merely most often occurs through cell proliferation.[one] This is considering a single cell with only one copy of the genome in the jail cell nucleus can perform biosynthesis and thus undergo cell growth at only one-half the rate of two cells. Hence, 2 cells abound (accumulate mass) at twice the rate of a single cell, and four cells abound at 4-times the rate of a single cell. This principle leads to an exponential increase of tissue growth charge per unit (mass aggregating) during cell proliferation, owing to the exponential increase in prison cell number.
Cell size depends on both cell growth and cell division, with a disproportionate increase in the rate of cell growth leading to production of larger cells and a disproportionate increase in the rate of jail cell division leading to product of many smaller cells. Cell proliferation typically involves counterbalanced cell growth and cell division rates that maintain a roughly constant prison cell size in the exponentially proliferating population of cells.
Some special cells tin grow to very large sizes via an unusual "endoreplication" jail cell cycle in which the genome is replicated during Southward-phase but at that place is no subsequent mitosis (Yard-stage) or cell division (cytokinesis). These large endoreplicating cells accept many copies of the genome, and then are highly polyploid.
Oocytes can be unusually large cells in species for which embryonic development takes place away from the female parent's body within an egg that is laid externally. The large size of some eggs tin can be accomplished either by pumping in cytosolic components from next cells through cytoplasmic bridges named band canals (Drosophila) or past internalisation of nutrient storage granules (yolk granules) past endocytosis (frogs).
Mechanisms of cell growth control [edit]
Cells tin can abound by increasing the overall rate of cellular biosynthesis such that production of biomolecules exceeds the overall rate of cellular degradation of biomolecules via the proteasome, lysosome or autophagy.
Biosynthesis of biomolecules is initiated past expression of genes which encode RNAs and/or proteins, including enzymes that catalyse synthesis of lipids and carbohydrates.
Individual genes are generally expressed via transcription into messenger RNA (mRNA) and translation into proteins, and the expression of each gene occurs to various different levels in a cell-type specific style (in response to gene regulatory networks).
To drive cell growth, the global rate of factor expression tin can be increased past enhancing the overall rate of transcription by RNA polymerase 2 (for active genes) or the overall rate of mRNA translation into poly peptide by increasing the abundance of ribosomes and tRNA, whose biogenesis depends on RNA polymerase I and RNA polymerase Three. The Myc transcription cistron is an case of a regulatory protein that tin can induce the overall activity of RNA polymerase I, RNA polymerase II and RNA polymerase III to bulldoze global transcription and translation and thereby cell growth.
In addition, the activity of individual ribosomes tin exist increased to boost the global efficiency of mRNA translation via regulation of translation initiation factors, including the 'translational elongation initiation cistron 4E' (eIF4E) complex, which binds to and caps the 5' end of mRNAs. The protein TOR, part of the TORC1 circuitous, is an important upstream regulator of translation initiation as well as ribosome biogenesis.[5] TOR is a serine/threonine kinase that can directly phosphorylate and inactivate a general inhibitor of eIF4E, named 4E-binding poly peptide (4E-BP), to promote translation efficiency. TOR likewise directly phosphorylates and activates the ribosomal poly peptide S6-kinase (S6K), which promotes ribosome biogenesis.
To inhibit cell growth, the global charge per unit of gene expression tin can exist decreased or the global rate of biomolecular degradation can be increased by increasing the rate of autophagy. TOR normally directly inhibits the part of the autophagy inducing kinase Atg1/ULK1. Thus, reducing TOR activity both reduces the global rate of translation and increases the extent of autophagy to reduce prison cell growth.
Cell growth regulation in animals [edit]
Many of the indicate molecules that command of cellular growth are chosen growth factors, many of which induce signal transduction via the PI3K/AKT/mTOR pathway, which includes upstream lipid kinase PI3K and the downstream serine/threonine poly peptide kinase Akt, which is able to activate some other protein kinase TOR, which promotes translation and inhibits autophagy to drive prison cell growth.
Nutrient availability influences production of growth factors of the Insulin/IGF-one family unit, which circulate as hormones in animals to actuate the PI3K/AKT/mTOR pathway in cells to promote TOR activity and so that when animals are well fed they will abound apace and when they are non able to receive sufficient nutrients they will reduce their growth rate.
In improver, the availability of amino acids to individual cells also directly promotes TOR activity, although this style of regulation is more important in single-celled organisms than in multicellular organisms such as animals that always maintain an abundance of amino acids in circulation.
Ane disputed theory proposes that many dissimilar mammalian cells undergo size-dependent transitions during the jail cell wheel. These transitions are controlled by the cyclin-dependent kinase Cdk1.[vi] Though the proteins that command Cdk1 are well understood, their connexion to mechanisms monitoring cell size remains elusive.
A postulated model for mammalian size control situates mass equally the driving forcefulness of the cell wheel. A prison cell is unable to grow to an abnormally large size because at a certain jail cell size or cell mass, the S phase is initiated. The S phase starts the sequence of events leading to mitosis and cytokinesis. A cell is unable to get too small considering the later prison cell cycle events, such as S, G2, and G, are delayed until mass increases sufficiently to begin S phase.[7]
Cell populations [edit]
Cell populations go through a particular blazon of exponential growth called doubling or prison cell proliferation. Thus, each generation of cells should be twice as numerous as the previous generation. However, the number of generations only gives a maximum figure every bit not all cells survive in each generation. Cells can reproduce in the stage of Mitosis, where they double and split into ii genetically equal cells.
Jail cell size [edit]
Cell size is highly variable among organisms, with some algae such as Caulerpa taxifolia existence a single prison cell several meters in length.[8] Establish cells are much larger than animal cells, and protists such as Paramecium tin can be 330 μm long, while a typical homo jail cell might exist x μm. How these cells "decide" how big they should be before dividing is an open question. Chemical gradients are known to exist partly responsible, and it is hypothesized that mechanical stress detection by cytoskeletal structures is involved. Piece of work on the topic generally requires an organism whose prison cell wheel is well-characterized.
Yeast cell size regulation [edit]
The human relationship betwixt cell size and prison cell division has been extensively studied in yeast. For some cells, there is a mechanism by which cell division is not initiated until a prison cell has reached a certain size. If the nutrient supply is restricted (after time t = 2 in the diagram, beneath), and the charge per unit of increment in jail cell size is slowed, the time period between cell divisions is increased.[nine] Yeast cell-size mutants were isolated that begin cell sectionalisation before reaching a normal/regular size (wee mutants).[10]

Effigy 1:Cell cycle and growth
Wee1 protein is a tyrosine kinase that normally phosphorylates the Cdc2 cell cycle regulatory protein (the homolog of CDK1 in humans), a cyclin-dependent kinase, on a tyrosine residue. Cdc2 drives entry into mitosis by phosphorylating a wide range of targets. This covalent modification of the molecular structure of Cdc2 inhibits the enzymatic activity of Cdc2 and prevents jail cell division. Wee1 acts to keep Cdc2 inactive during early G2 when cells are still small. When cells take reached sufficient size during G2, the phosphatase Cdc25 removes the inhibitory phosphorylation, and thus activates Cdc2 to let mitotic entry. A balance of Wee1 and Cdc25 action with changes in cell size is coordinated past the mitotic entry command system. It has been shown in Wee1 mutants, cells with weakened Wee1 activeness, that Cdc2 becomes active when the jail cell is smaller. Thus, mitosis occurs before the yeast attain their normal size. This suggests that cell sectionalisation may be regulated in role by dilution of Wee1 protein in cells every bit they grow larger.
Linking Cdr2 to Wee1 [edit]
The poly peptide kinase Cdr2 (which negatively regulates Wee1) and the Cdr2-related kinase Cdr1 (which directly phosphorylates and inhibits Wee1 in vitro)[11] are localized to a band of cortical nodes in the center of interphase cells. Later entry into mitosis, cytokinesis factors such as myosin Ii are recruited to like nodes; these nodes eventually condense to form the cytokinetic ring.[12] A previously uncharacterized protein, Blt1, was establish to colocalize with Cdr2 in the medial interphase nodes. Blt1 knockout cells had increased length at segmentation, which is consistent with a filibuster in mitotic entry. This finding connects a physical location, a band of cortical nodes, with factors that have been shown to directly regulate mitotic entry, namely Cdr1, Cdr2, and Blt1.
Farther experimentation with GFP-tagged proteins and mutant proteins indicates that the medial cortical nodes are formed by the ordered, Cdr2-dependent associates of multiple interacting proteins during interphase. Cdr2 is at the top of this hierarchy and works upstream of Cdr1 and Blt1.[13] Mitosis is promoted by the negative regulation of Wee1 by Cdr2. It has also been shown that Cdr2 recruits Wee1 to the medial cortical node. The machinery of this recruitment has yet to exist discovered. A Cdr2 kinase mutant, which is able to localize properly despite a loss of function in phosphorylation, disrupts the recruitment of Wee1 to the medial cortex and delays entry into mitosis. Thus, Wee1 localizes with its inhibitory network, which demonstrates that mitosis is controlled through Cdr2-dependent negative regulation of Wee1 at the medial cortical nodes.[13]
Cell polarity factors [edit]
Cell polarity factors positioned at the jail cell tips provide spatial cues to limit Cdr2 distribution to the cell middle. In fission yeast Schizosaccharomyces pombe (Due south. Pombe), cells divide at a defined, reproducible size during mitosis considering of the regulated activity of Cdk1.[14] The jail cell polarity protein kinase Pom1, a member of the dual-specificity tyrosine-phosphorylation regulated kinase (DYRK) family unit of kinases, localizes to cell ends. In Pom1 knockout cells, Cdr2 was no longer restricted to the cell middle, but was seen diffusely through half of the prison cell. From this data it becomes credible that Pom1 provides inhibitory signals that confine Cdr2 to the middle of the prison cell. It has been farther shown that Pom1-dependent signals lead to the phosphorylation of Cdr2. Pom1 knockout cells were besides shown to divide at a smaller size than wild-type, which indicates a premature entry into mitosis.[13]
Pom1 forms polar gradients that peak at jail cell ends, which shows a direct link between size control factors and a specific physical location in the cell.[15] As a jail cell grows in size, a slope in Pom1 grows. When cells are small, Pom1 is spread diffusely throughout the cell body. Every bit the cell increases in size, Pom1 concentration decreases in the middle and becomes concentrated at cell ends. Small cells in early on G2 which contain sufficient levels of Pom1 in the entirety of the cell have inactive Cdr2 and cannot enter mitosis. It is not until the cells grow into late G2, when Pom1 is confined to the cell ends that Cdr2 in the medial cortical nodes is activated and able to start the inhibition of Wee1. This finding shows how cell size plays a straight role in regulating the kickoff of mitosis. In this model, Pom1 acts as a molecular link betwixt prison cell growth and mitotic entry through a Cdr2-Cdr1-Wee1-Cdk1 pathway.[13] The Pom1 polar gradient successfully relays data about jail cell size and geometry to the Cdk1 regulatory system. Through this slope, the cell ensures it has reached a divers, sufficient size to enter mitosis.
Other experimental systems for the study of jail cell size regulation [edit]
One mutual means to produce very large cells is by cell fusion to form syncytia. For example, very long (several inches) skeletal muscle cells are formed by fusion of thousands of myocytes. Genetic studies of the fruit fly Drosophila have revealed several genes that are required for the formation of multinucleated muscle cells past fusion of myoblasts.[16] Some of the key proteins are of import for prison cell adhesion between myocytes and some are involved in adhesion-dependent cell-to-jail cell signal transduction that allows for a cascade of jail cell fusion events.
Increases in the size of plant cells are complicated past the fact that nigh all plant cells are inside of a solid cell wall. Under the influence of certain institute hormones the cell wall can be remodeled, assuasive for increases in cell size that are important for the growth of some plant tissues.
Near unicellular organisms are microscopic in size, but there are some behemothic bacteria and protozoa that are visible to the naked eye. (Run into Tabular array of jail cell sizes—Dumbo populations of a behemothic sulfur bacterium in Namibian shelf sediments[17]—Large protists of the genus Anarchy, closely related to the genus Amoeba.)
In the rod-shaped bacteria E. coli, Caulobacter crescentus and B. subtilis cell size is controlled by a simple mechanisms in which cell division occurs after a constant volume has been added since the previous segmentation.[18] [19] Past always growing by the same amount, cells born smaller or larger than average naturally converge to an average size equivalent to the amount added during each generation.
Jail cell division [edit]
Prison cell reproduction is asexual. For most of the constituents of the cell, growth is a steady, continuous process, interrupted merely briefly at Thou phase when the nucleus and then the cell divide in two.
The process of prison cell division, called cell cycle, has four major parts called phases. The get-go part, called G1 phase is marked by synthesis of various enzymes that are required for DNA replication. The second part of the prison cell bicycle is the S stage, where DNA replication produces two identical sets of chromosomes. The third role is the G2 stage in which a significant protein synthesis occurs, mainly involving the production of microtubules that are required during the process of division, called mitosis. The quaternary phase, M phase, consists of nuclear division (karyokinesis) and cytoplasmic partitioning (cytokinesis), accompanied by the formation of a new cell membrane. This is the physical segmentation of "mother" and "daughter" cells. The Grand phase has been broken downwardly into several distinct phases, sequentially known equally prophase, prometaphase, metaphase, anaphase and telophase leading to cytokinesis.
Prison cell division is more circuitous in eukaryotes than in other organisms. Prokaryotic cells such every bit bacterial cells reproduce by binary fission, a process that includes DNA replication, chromosome segregation, and cytokinesis. Eukaryotic cell division either involves mitosis or a more than complex process called meiosis. Mitosis and meiosis are sometimes chosen the ii "nuclear division" processes. Binary fission is similar to eukaryote prison cell reproduction that involves mitosis. Both lead to the production of two daughter cells with the same number of chromosomes as the parental prison cell. Meiosis is used for a special cell reproduction process of diploid organisms. Information technology produces 4 special daughter cells (gametes) which take one-half the normal cellular amount of Deoxyribonucleic acid. A male and a female gamete can then combine to produce a zygote, a cell which once again has the normal corporeality of chromosomes.
The balance of this article is a comparison of the principal features of the three types of cell reproduction that either involve binary fission, mitosis, or meiosis. The diagram below depicts the similarities and differences of these three types of jail cell reproduction.

Comparison of the three types of cell division [edit]
The DNA content of a cell is duplicated at the start of the cell reproduction process. Prior to Dna replication, the DNA content of a jail cell tin be represented as the amount Z (the cell has Z chromosomes). After the DNA replication process, the amount of Dna in the cell is 2Z (multiplication: two 10 Z = 2Z). During Binary fission and mitosis the duplicated DNA content of the reproducing parental cell is separated into 2 equal halves that are destined to terminate upwards in the two daughter cells. The final part of the jail cell reproduction process is cell division, when daughter cells physically split apart from a parental cell. During meiosis, at that place are two cell sectionalization steps that together produce the iv daughter cells.
After the completion of binary fission or jail cell reproduction involving mitosis, each girl jail cell has the same amount of DNA (Z) as what the parental cell had before it replicated its Dna. These two types of prison cell reproduction produced two daughter cells that take the same number of chromosomes as the parental cell. Chromosomes duplicate prior to jail cell partitioning when forming new pare cells for reproduction. Afterwards meiotic jail cell reproduction the four daughter cells take half the number of chromosomes that the parental cell originally had. This is the haploid amount of DNA, often symbolized every bit N. Meiosis is used by diploid organisms to produce haploid gametes. In a diploid organism such as the human organism, most cells of the body have the diploid corporeality of Deoxyribonucleic acid, 2N. Using this annotation for counting chromosomes we say that homo somatic cells have 46 chromosomes (2N = 46) while man sperm and eggs have 23 chromosomes (North = 23). Humans have 23 distinct types of chromosomes, the 22 autosomes and the special category of sex chromosomes. In that location are ii singled-out sex chromosomes, the Ten chromosome and the Y chromosome. A diploid human being prison cell has 23 chromosomes from that person's father and 23 from the female parent. That is, your body has two copies of human being chromosome number 2, ane from each of your parents.

Immediately after Deoxyribonucleic acid replication a human cell will take 46 "double chromosomes". In each double chromosome there are 2 copies of that chromosome'south Deoxyribonucleic acid molecule. During mitosis the double chromosomes are split to produce 92 "unmarried chromosomes", half of which go into each girl cell. During meiosis, there are two chromosome separation steps which assure that each of the iv daughter cells gets one copy of each of the 23 types of chromosome.
Sexual reproduction [edit]
Though jail cell reproduction that uses mitosis tin can reproduce eukaryotic cells, eukaryotes bother with the more complicated process of meiosis because sexual reproduction such as meiosis confers a selective reward. Notice that when meiosis starts, the two copies of sister chromatids number 2 are adjacent to each other. During this fourth dimension, in that location can be genetic recombination events. Information from the chromosome 2 DNA gained from one parent (red) will transfer over to the chromosome two Deoxyribonucleic acid molecule that was received from the other parent (greenish). Detect that in mitosis the two copies of chromosome number ii do not interact. Recombination of genetic information between homologous chromosomes during meiosis is a process for repairing DNA amercement. This procedure can besides produce new combinations of genes, some of which may be adaptively beneficial and influence the course of development. However, in organisms with more than one gear up of chromosomes at the chief life bike stage, sexual practice may also provide an reward because, under random mating, it produces homozygotes and heterozygotes according to the Hardy–Weinberg ratio.
Disorders [edit]
A series of growth disorders can occur at the cellular level and these consequently underpin much of the subsequent course in cancer, in which a group of cells display uncontrolled growth and division beyond the normal limits, invasion (intrusion on and destruction of adjacent tissues), and sometimes metastasis (spread to other locations in the body via lymph or claret). Several key determinants of jail cell growth, like ploidy and the regulation of cellular metabolism, are commonly disrupted in tumors.[twenty] Therefore, heterogenous cell growth and pleomorphism is 1 of the earliest hallmarks of cancer progression.[21] [22] Despite the prevalence of pleomorphism in human pathology, its role in affliction progression is unclear. In epithelial tissues, pleomorphism in cellular size can induce packing defects and disperse aberrant cells.[23] Merely the result of singular jail cell growth in other animate being tissues is unknown.
Measurement methods [edit]
The cell growth can be detected by a variety of methods. The cell size growth can be visualized by microscopy, using suitable stains. But the increase of cells number is commonly more significant. It tin exist measured by transmission counting of cells under microscopy observation, using the dye exclusion method (i.eastward. trypan blueish) to count only feasible cells. Less fastidious, scalable, methods include the utilise of cytometers, while flow cytometry allows combining cell counts ('events') with other specific parameters: fluorescent probes for membranes, cytoplasm or nuclei permit distinguishing dead/viable cells, cell types, cell differentiation, expression of a biomarker such equally Ki67.
Abreast the increasing number of cells, 1 can be assessed regarding the metabolic activity growth, that is, the CFDA and calcein-AM measure out (fluorimetrically) not only the membrane functionality (dye memory), but also the functionality of cytoplasmic enzymes (esterases). The MTT assays (colorimetric) and the resazurin assay (fluorimetric) dose the mitochondrial redox potential.
All these assays may correlate well, or non, depending on cell growth conditions and desired aspects (activity, proliferation). The task is fifty-fifty more than complicated with populations of different cells, furthermore when combining cell growth interferences or toxicity.
Run across also [edit]
- Bacterial growth
References [edit]
- ^ a b c Conlon, Ian; Raff, Martin (1999). "Size Control in Animal Development". Cell. 96 (2): 235–244. doi:ten.1016/S0092-8674(00)80563-2. ISSN 0092-8674. PMID 9988218. S2CID 15738174.
- ^ Grewal, Savraj South; Edgar, Bruce A (2003). "Controlling prison cell division in yeast and animals: does size affair?". Journal of Biology. 2 (1): 5. doi:10.1186/1475-4924-2-5. ISSN 1475-4924. PMC156596. PMID 12733996.
- ^ Neufeld, Thomas P; de la Cruz, Aida Flor A; Johnston, Laura A; Edgar, Bruce A (1998). "Coordination of Growth and Cell Division in the Drosophila Wing". Cell. 93 (7): 1183–1193. doi:10.1016/S0092-8674(00)81462-ii. ISSN 0092-8674. PMID 9657151. S2CID 14608744.
- ^ Thompson, Barry J. (2010). "Developmental control of prison cell growth and division in Drosophila". Current Opinion in Cell Biological science. 22 (6): 788–794. doi:x.1016/j.ceb.2010.08.018. PMID 20833011.
- ^ Hafen, Due east. (2004). "Coaction Between Growth Factor and Nutrient Signaling: Lessons from Drosophila TOR". TOR. Electric current Topics in Microbiology and Immunology. Vol. 279. pp. 153–167. doi:10.1007/978-3-642-18930-2_10. ISBN978-three-642-62360-8. ISSN 0070-217X. PMID 14560957.
- ^ Mitchison JM (2003). "Growth during the cell cycle". Int. Rev. Cytol. International Review of Cytology. 226: 165–258. doi:10.1016/S0074-7696(03)01004-0. ISBN978-0-12-364630-9. PMID 12921238.
- ^ Cooper, Stephen (2004). "Control and maintenance of mammalian cell size". BMC Prison cell Biology. 5 (ane): 35. doi:x.1186/1471-2121-v-35. PMC524481. PMID 15456512.
- ^ Peplow, Mark (23 March 2005). "Algae create mucilage to repair cell damage". Nature.com . Retrieved 4 July 2016.
- ^ Slavov Due north.; Botstein D. (June 2011). "Coupling among Growth Charge per unit Response, Metabolic Cycle and Cell Partitioning Bicycle in Yeast". Molecular Biology of the Jail cell. 22 (12): 1997–2009. doi:10.1091/mbc.E11-02-0132. PMC3113766. PMID 21525243.
- ^ Wee1 mutants of South. pombe have modest cell size and the homologous proteins in humans also regulate cell entry into mitosis; in Lodish HF, Berk A, Zipursky LS, Matsudaira P, et al., eds. (2000). Molecular cell biology (4th ed.). New York: W.H. Freeman. ISBN978-0-7167-3136-8.
- ^ Wu L, Russell P (June 1993). "Nim1 kinase promotes mitosis by inactivating Wee1 tyrosine kinase". Nature. 363 (6431): 738–41. Bibcode:1993Natur.363..738W. doi:10.1038/363738a0. PMID 8515818. S2CID 4320080.
- ^ Wu JQ, Kuhn JR, Kovar DR, Pollard TD (November 2003). "Spatial and temporal pathway for assembly and constriction of the contractile ring in fission yeast cytokinesis". Dev. Cell. 5 (five): 723–34. doi:10.1016/S1534-5807(03)00324-1. PMID 14602073.
- ^ a b c d Moseley JB, Mayeux A, Paoletti A, Nurse P (June 2009). "A spatial gradient coordinates cell size and mitotic entry in fission yeast". Nature. 459 (7248): 857–sixty. Bibcode:2009Natur.459..857M. doi:x.1038/nature08074. PMID 19474789. S2CID 4330336.
- ^ Rupes I (September 2002). "Checking cell size in yeast". Trends Genet. xviii (9): 479–85. doi:10.1016/S0168-9525(02)02745-2. PMID 12175809.
- ^ Padte NN, Martin SG, Howard M, Chang F (December 2006). "The cell-end cistron pom1p inhibits mid1p in specification of the cell division aeroplane in fission yeast". Curr. Biol. 16 (24): 2480–7. doi:10.1016/j.cub.2006.11.024. PMID 17140794.
- ^ Menon SD, Osman Z, Chenchill G, Chia W (June 2005). "A positive feedback loop between Dumbfounded and Rolling pebbles leads to myotube enlargement in Drosophila". J. Cell Biol. 169 (6): 909–20. doi:ten.1083/jcb.200501126. PMC2171639. PMID 15955848.
- ^ Schulz HN, Brinkhoff T, Ferdelman TG, Mariné MH, Teske A, Jorgensen BB (April 1999). "Dumbo populations of a behemothic sulfur bacterium in Namibian shelf sediments". Scientific discipline. 284 (5413): 493–five. Bibcode:1999Sci...284..493S. doi:ten.1126/science.284.5413.493. PMID 10205058. S2CID 32571118.
- ^ Taheri-Araghi, Due south; Bradde, S; Sauls, J. T.; Hill, N. Due south.; Levin, P. A.; Paulsson, J; Vergassola, Grand; Jun, South (February 2015). "Cell-size command and homeostasis in bacteria". Current Biological science. 25 (3): 385–391. doi:10.1016/j.cub.2014.12.009. PMC4323405. PMID 25544609.
- ^ Campos, One thousand; Surovtsev, I. V.; Kato, S; Paintdakhi, A; Beltran, B; Ebmeier, S. E.; Jacobs-Wagner, C (Dec 2014). "A constant size extension drives bacterial cell size homeostasis". Jail cell. 159 (6): 1433–1446. doi:10.1016/j.cell.2014.11.022. PMC4258233. PMID 25480302.
- ^ Schmoller, Kurt M.; Skotheim, January Grand. (December 2015). "The Biosynthetic Footing of Cell Size Control". Trends Cell Biol. 25 (12): 793–802. doi:10.1016/j.tcb.2015.x.006. PMC6773270. PMID 26573465.
- ^ Travis, W.D.; Brambilla, B.; Burke, A.P; Marx, A.; Nicholson, A.G. (2015). WHO Classification of Tumours of the Lung, Pleura, Thymus and Heart. Lyon: International Bureau for Inquiry on Cancer. ISBN978-92-832-2436-5.
- ^ El-Naggar, A.K.; Chan, J.C.K.; Grandis, J.R.; Takata, T.; Slootweg, P.J. (2017-01-23). WHO Classification of Head and Neck Tumours. Lyon: International Agency for Enquiry on Cancer. ISBN978-92-832-2438-9. Archived from the original on 2019-10-31. Retrieved 2019-10-31 .
- ^ Ramanathan, Subramanian P.; Krajnc, Matej; Gibson, Matthew C. (October 2019). "Jail cell-Size Pleomorphism Drives Aberrant Clone Dispersal in Proliferating Epithelia". Developmental Cell. 51 (1): 49–61.e4. doi:x.1016/j.devcel.2019.08.005. PMC6903429. PMID 31495693.
Books [edit]
- Morgan, David O. (2007). The cell cycle: principles of control. London: Sunderland, Mass. ISBN978-0-9539181-ii-vi.
External links [edit]
- A comparing of generational and exponential models of cell population growth
- Local Growth in an Array of Disks Wolfram Demonstrations Project
0 Response to "Cells Operate in an Order Fashion Under the Influence of"
Post a Comment